

Primerament la recerca de fauna subterrània és localitzà a les cavitats naturals, aviat és comprovà que igualment colonitzaven altres hipogeus, més recentment la investigació ha anat trobant un altre habitat colonitzat per una part d'aquesta fauna, el MSS (col·luvions, tarteres, pedregars, sols, etc) i ara se'ns presenta un nou terreny de recerca en el medi al·luvial. Els autors ens presenten la seva recerca experimental, material utilitzat i primeres conclusions.
Ortuño VM, Gilgado JD, Jiménez-Valverde A, Sendra A, Pérez-Suárez G, et al.
(2013) The “Alluvial Mesovoid Shallow Substratum”, a New Subterranean Habitat
PLoS ONE 8(10): e76311. doi:10.1371/journal.pone.0076311
El treball complert el podeu trobar a:
http://www.plosone.org/article/info%3Adoi/10.1371/journal.pone.0076311
Tot seguit trobareu una selecció dels continguts d'aquest interessant article:
Abstract
In this paper we describe a new type of subterranean habitat associated
with dry watercourses in the Eastern Iberian Peninsula, the “Alluvial Mesovoid Shallow
Substratum” (alluvial MSS). Historical observations and data from field
sampling specially designed to study MSS fauna in the streambeds of temporary watercourses
support the description of this new habitat. To conduct the sampling, 16 subterranean
sampling devices were placed in a region of Eastern Spain. The traps were
operated for 12 months and temperature and relative humidity data were recorded
to characterise the habitat. A large number of species was captured, many of
which belonged to the arthropod group, with marked hygrophilous, geophilic, lucifugous
and mesothermal habits. In addition, there was also a substantial number of species
showing markedly ripicolous traits. The results confirm that the network of spaces
which forms in alluvial deposits of temporary watercourses merits the category
of habitat, and here we propose the name of “alluvial MSS”. The “alluvial MSS” may
be covered or not by a layer of soil, is extremely damp, provides a buffer against
above ground temperatures and is aphotic. In addition, compared to other types of
MSS, it is a very unstable habitat. It is possible that the “alluvial MSS” may
be found in other areas of the world with strongly seasonal climatic regimes, and
could play an important role as a biogeographic corridor and as a refuge from
climatic changes.
For nearly
eight decades since the appearance of biospeleology as a scientific discipline
following the publication of the study by Racovitza [1],
classic authors such as Jeannel [2,3],
Vandel [4]
or Ginet & Decou [5]
have been drawing attention to the extent of the subterranean domain in cracks
and fissures beneath the soil surface (microcaverns and mesocaverns sensu
Howarth [6]),
besides the caves accessible to humans. However, it was not until the 1980s
that the studies led by Christian Juberthie [7,8]
and the observations carried out by Shun-Ichi Uéno [9,10]
revealed the extent of the subterranean domain beneath the karst surface
shallow, designated Milieu Souterrain Superficiel in French, and later
translated to English as Mesovoid Shallow Substratum, abbreviated to MSS. It
forms a unique habitat with its own fauna, but can also play the role of an
ecotone, in which both epigean and truly subterranean, troglobiont life forms
thrive. A continuum of life is established from the surface soil, through the
different edaphic horizons and the MSS, and finally, in the deep subterranean
environment, as reported in a study by Gers [11].
A basic
condition for the existence of the MSS is the presence of rocky deposits that
have generated - and preserved - subterranean spaces which life forms can
inhabit. There are many diverse forms of the MSS habitat, which arise as the
result of several abiotic and biotic factors [12].
To date, three basic types of MSS have been distinguished:
1): Colluvial or slope MSS
This
is formed on sloping ground [7],
and is the result of the erosion and deposition of rock fragments produced by
the mechanical fragmentation of rock walls or the erosive action of continental
glaciers. The nature of the rock can be diverse, including calcareous,
siliceous and even volcanic rocks. These rocky deposits can be covered by
evolving soil (with edaphic horizons) of different thicknesses which often
supports dense plant cover. In other cases, the upper level of the MSS is bare,
appearing as mountain scree.
2): Bedrock MSS
This
is formed by the weathering of much of the bedrock [13],
a gradual process that occurs almost simultaneously with the formation of
edaphic horizons which seal it off from the external environment. This type of
MSS is found in valley bottoms or areas with little or no slope. The nature of
the rock may be very diverse, although it is more likely to occur in rocks that
are easily altered.
3): Volcanic MSS
This
is the result of the accumulation of volcanic material [14].
From formation onwards, it contains many interconnected fissures forming a
tangled web of micro-spaces, which over time become isolated from the outside
due to the development of an edaphic environment [15].
This type of MSS is more ephemeral than others, since deposits of volcanic
material are more susceptible to weathering than those composed of more compact
rocks such as sedimentary, plutonic or metamorphic rocks.
In this
paper, we present a proposal supported by experimental and theoretical
arguments for a new type of MSS which we have named “alluvial MSS”, and which corresponds
to the crevices (unfilled spaces) which lie beneath the streambeds of temporary
watercourses.
The existence
of hypogean species in watercourses was already known from occasional citations
[16,17],
but it was the search for Thalassophilus breuili Jeannel, 1926, endemic to Alicante,
that triggered the study which led us to propose a new type of MSS habitat, the
“alluvial MSS”. Taking into account that this species was so rare in caves (only
three specimens from two caves are known [18,19])
and that the third time it was cited, it was on the banks of a temporary watercourse
(the Guadalest River) in Altea (Alicante, Eastern Spain) [17],
this suggested the possibility that this species might have been swept away by water
from a nearby MSS, and that it might be found in this latter habitat [20].
Given the extraordinary irregularity of the hydrological regime of watercourses
in the province of Alicante, it was thought that perhaps T. breuili might live in
the substratum of these streambeds. Pursuing this lead, one of the authors
(V.M.O.) installed 4 pitfall traps, buried 50-70 cm deep, in the streambed of the
watercourse known as the “Barranco de la Cueva de los Corrales” (Xixona, Alicante,
Spain). These traps were operated for 4 months (June-September, 2009), collecting
a considerable number of arthropods, which abounded in the interstices of the substratum
of the watercourse. Although T. breuili was not found, it is worth noting the capture
of more than a hundred Speonemadus sp., Coleoptera Leiodidae typical of large subterranean
cavities and MSS [21,22].
All these data prompted the launch of a research project aimed, among other
objectives, at prospecting and studying the subterranean spaces located among the
alluvial deposits of dry watercourses in the province of Alicante. These watercourses
are dry for most of the year, carrying water only during very sporadic periods of
heavy rainfall. However, this subterranean environment remained damp throughout
the year and a priori, its structure, comprising stony debris with an extensive
network of crevices, could provide a refuge for terrestrial stenohygrobic
fauna.
Study area
The
geographic area selected for our study encompassed the eastern end of the karst
reliefs of the Prebetic range, from the centre and north of the province of
Alicante to the south of the province of Valencia (Eastern Spain), and was
chosen because there are numerous temporary watercourses in the area, thanks to
a series of geological, geographical and climatic factors. In the local
toponymy, these types of watercourse are termed “rambla” (low, wide channel)
and “barranco” or “barranc” (tall, narrow channel), words which reflect
cultural and linguistic factors [23,24]
rather than a true hydrographic characterisation. Regardless, these dry
watercourses are usually dry for most of the year, only carrying water for an
average of ten days a year [24,25].
The study
area was divided into first-order river basins (Figure
1), following the criterion of IDEJúcar [26],
and subsequently, basins in the northern half, the wettest, most mountainous area,
were selected. A preliminary selection of promising sites for sampling was made
using Sigpac [27]
and Google, Earth [28].
Of the 42 sites selected a priori, 16 were considered eligible (Figure
1; Table
1) because the watercourses they contained were dry for most of the year and
had thick alluvial deposits with numerous subterranean spaces. The other sites were
rejected because they did not present either of the two features mentioned. Many
of them overlay inappropriate lithologies, as was the case with the marly
series of the Tap [29]
which constitute physical barriers to the movement of subterranean fauna [30].

Description of a New Subterranean Habitat: The
“alluvial MSS”
For
the most part, the fauna communities found in the alluvial substratum consisted
of species belonging to four groups with well-defined ecophysiological
characteristics, being geophilic, hygrophilous, mesothermal and/or lucifugous
species. These species exhibit a geophilic nature, inhabiting environments
closely associated with soil (edaphic horizons or different epiedaphic habitats
and biotopes). They can be either totally geophilic, i.e. their entire life
cycle occurs in these habitats/biotopes, or partially geophilic, in which only
the larval stages inhabit this type of environment, as in the case of
Lampyridae species of the genera Nyctophila and Lamprohiza, both frequently
observed in the alluvial substratum. The second ecophysiological characteristic
is the strongly hygrophilous nature of the species which were well represented
in the alluvial substratum. In areas such as that studied, with a Mediterranean
climate, these highly hygrophilous species are extremely rare or absent in
edaphic/epiedaphic environments. The third characteristic is the mesothermal
nature of many of these species, a circumstance that forces them to seek refuge
in favourable environments which mitigate the sudden daily and/or seasonal
changes in temperature. The last ecophysiological characteristic was the
lucifugous nature which, however, only a portion of the ensemble of alluvial
substratum fauna exhibited. This characteristic may be expressed to a lesser or
greater degree, and thus some species may show a preference for shaded
environments (sciophily) whilst others thrive in aphotic environments (a
diversity of subterranean environments).
Different
species presented different combinations of these characteristics. There were
strongly hygrophilous, lucifugous and mesothermal species, all found in caves
and other subterranean spaces, such as species belonging to the lineage Trechus
martinezi, Lepthyphantes spp. and certain Isopoda Oniscidea, among others, which
are considered troglobiont forms. Then there were strongly hygrophilous,
mesothermal and sciophilic species (not exclusive to aphotic environments),
such as Dysdera spp., Lithobius spp., Porotachys bisulcatus, Ocys harpaloides,
Platyderus spp., Speonemadus spp. and some Oniscidea Isopoda, among others,
which constitute an ensemble of species which are typically troglophilic. There
were also moderately hygrophilous, mesothermal and sciophilic species which may
sometimes behave like true troglophiles, such as Dicranolasma soerensenii and
Petaloptila spp., and lastly, there were species which were markedly
hygrophilous, may or may not be mesothermal and were not lucifugous, as is the
case of fauna exhibiting ripicolous behaviour such as Pardosa spp. and some of
the Carabidae Bembidiinae species, among others.
Thus,
as happens in caves, the alluvial substratum constitutes a habitat occupied by
fauna which is exclusively hypogean (troglobionts), frequently hypogean
(troglophiles), and where numerous edaphic species occur (Acari and Collembola,
among others) to a different degree, in addition to ripicolous and trogloxene
fauna. The latter group consisted of species with a small number of individuals
belonging to different orders (Diptera and Hymenoptera, among others), for
which living conditions in the substratum are not optimal. This group of
specimens reached the substratum by accident, either actively, as part of their
daily scavenging activities in the edaphic/epiedaphic environment, or
passively, usually as a result of hydrochory and also zoochory. Despite being
considered species which are not adapted to this environment, they make a vital
contribution to the flow of energy in these subterranean habitats [11,58].
Therefore, there is fauna with different ecological roles: some of them shared
with caves and other MSSs (troglobionts and trogloxenes), which characterizes
this habitat as a true MSS; some others are typical of soil (edaphobionts),
which are also frequent in other MSS [59];
epigean fauna (trogloxenes), due to its proximity to the surface and the
conditions of temperature and humidity; and finally ripicolous fauna, whose
presence would be particularly characteristic of this habitat.

We
propose naming these subterranean spaces located in the alluvial substratum
“alluvial MSS”, a hypogean habitat composed of cracks and crevices in the
substratum that form among the gravel and variously sized pebbles that
constitute the alluvial deposits of dry watercourses (Figure
4). To date, all approaches to the study and recognition of fauna in the
substratum of watercourses have focused on the hyporheic zone and,
collaterally, with certain adjacent biotopes, some of which might fall under
the classification of MSS. The hyporheic zone [60,61]
is limited to the substratum of active watercourses, through the interstices of
which water circulates and provides a habitat for aquatic species, many of
which can be described as stygobionts. In addition, the dynamics of the recent
past of these rivers has led to the formation of river terraces beneath which
groundwater circulates, constituting aquifers (phreatic biotopes) that may be
in contact with the waters of rivers and streams; this forms another type of
subterranean water environment [61,]
where invertebrate stygobionts are also frequently present. With regard to the MSS
that is subject to a fluvial influence and whose origin is unrelated to the dynamics
of river currents, Uéno [62]
revealed the importance of colluvial deposits and slopes that are more or less directly
in contact with the waters of rivers with a moderate current.

The “alluvial
MSS” appears in dry watercourses, which appear as scars on the terrain, through
which water only flows in times of flood, creating torrential type hydrological
regimes. This hydrological dynamic is associated with the Mediterranean climate
characteristic of the area, in which maximum rainfall occurs in autumn with a
smaller peak in spring, whilst the summer is characterised by a very marked
minimum rainfall accompanied by an increase in temperature [24].
Floods occur after irregular, heavy rainfall which starts suddenly and
sometimes violently [23,].
However, there are many types of flood, of a diverse nature and intensity [64].
It is of interest to note that those known as flash floods [65]
whilst not the only type to occur in the areas studied, have a very significant
effect since they can even push solid blocks of a diameter of up to twice the
height of the water [24,].
These phenomena may be due to different causes, such as summer convective
storms, weather fronts and orographic precipitation [67],
and often also to the phenomenon known as “cut-off low” [68-70],
responsible for numerous catastrophic episodes of flooding.
Since
the watercourses in these areas are dry for most of the year, the fissures in
the substratum are not permanently waterlogged. However, they retained
sufficient water even when water levels were at their lowest for the stones
removed during the installation of the SSDs to be damp (unpublished data), and
RHmean levels of 80% and RHmin levels of 46.5% were recorded for some of the
SSDs (Table
1, Barranco de la Cueva de los Corrales). This circumstance facilitates the
survival of plants growing in the streambed, provides a refuge for hygrophilous
terrestrial fauna and supports the presence of both exclusively and frequently
hypogean species, rendering this set of spaces among the stones of alluvial
debris a new type of MSS.
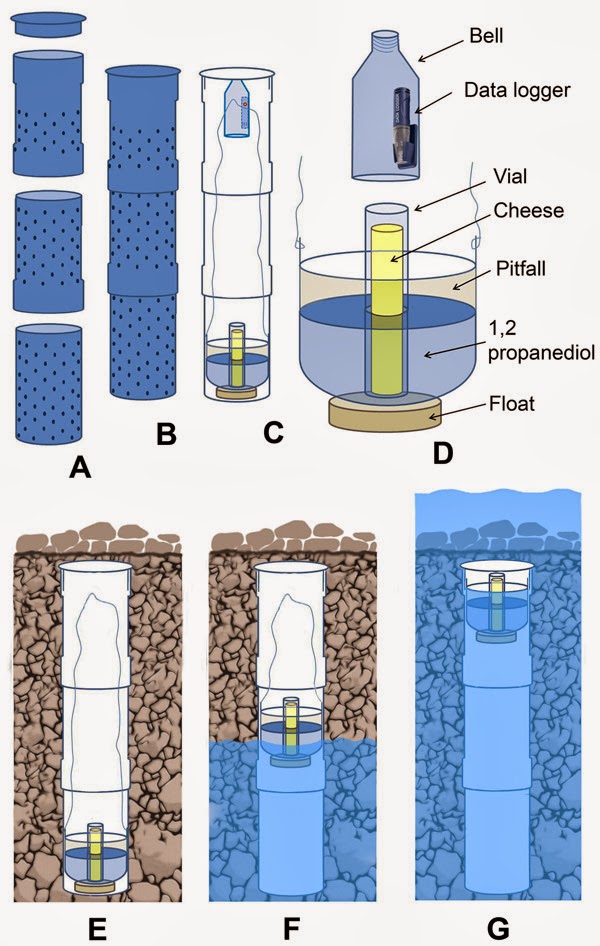
Formation and structure of the “alluvial MSS”
No
single factor determines the presence of temporary watercourses in the Eastern
Iberian Peninsula. In other regions of the Mediterranean, the presence of
temporary watercourses has been described as the result of a synergy of
geological, geomorphological and climatic factors [71].
However, in the case of the study area, there is no clear relationship between
the presence of such courses and a given geological substrate or a particular
rainfall pattern [23].
Furthermore, it should be noted that not all of the Eastern Iberian
watercourses are subject to a seasonal regime, as some have a constant flow of
water throughout the annual hydrological cycle, either along the entire course
or only a part of it, in which groundwater plays an important role in
maintaining a base flow [72].
It should also be noted that the presence of dry watercourses is not a feature
unique to the Iberian Peninsula. Rather, they appear in numerous regions of the
world with very different climates and geology, receiving different names: the
“Chapp” in the Gobi, “Laagate” in the Kalahari, “Wadi” in the Maghreb, “Donga”
in South Africa, “Nullah” in India, “Fiumare” in Italy, “Arroyo” in the U.S.A.
and Mexico, and “Dry Valley” in England [24,].
With
regard to the dry watercourses studied, these are known to have a considerable carrying
capacity and thus it is very common to see large boulders, pebbles, gravel and
sand on the streambed, forming a very heterogeneous group of rocks from
different sources [72].
The loss of the water’s inertia results in the rapid and massive sedimentation
of the transported materials [74].
This circumstance converts the lower reaches of these dry watercourses (the
low, wide channel known as the “rambla”) into areas where “alluvial MSS” is
unlikely to exist because the interstices between the boulders are generally
filled (Figure
5a, b). Our field work revealed the existence of “alluvial MSS” in the
upper and middle reaches of these watercourses, where the alluvial debris
formed by calcareous stones contained a network of empty spaces which had not
been filled by sediments, allowing the movement of fauna between these
interstices (Figure
6). However, we cannot assert that dry watercourses in different regions of
the world behave in the same way.
Although
the “alluvial MSS” is not covered by true soil, it is noteworthy that it may have
some degree of insulation from the surface as a result of one phenomenon which occurs
with relative frequency in these watercourses. This is the phenomenon known as
“armouring” [24,],
which occurs when flooding ceases and the carrying capacity of the watercourse
decreases. At this point, the coarser materials are deposited, forming
protective films or “armour”, and the finest materials filter between and
rearrange the interstices. Hence, a thin surface layer of silt, sand and gravel
is created between the larger stones close to the surface, which contributes to
insulating the MSS to a certain extent (Figure
4b, c, d).
In
addition, the vegetation that grows not only on the margins of these dry
watercourses but also in their channels, plays an important geomorphological
role, proffering mechanical resistance to the water and anchoring the alluvial
deposits with their roots. Hence, they provide the “alluvial MSS” with greater
structural stability against the drag force of torrential waters and favour
sedimentation of the particles carried by the water, thus helping to increase
the thickness of these deposits [76].
The dry watercourses studied were mostly colonised by oleander formations,
shrubs which are characteristic of dry Mediterranean environments and are
adapted to withstand both hot drought and the force of floodwaters. Although
the oleander (Nerium oleander L.) predominated in these formations, it was
often accompanied by various thorny bushes (mostly of the genera Rubus L, Rosa
L. and Crataegus L.) and by nitrophilous plants whose growth was favoured by
the contribution of organic detritus periodically deposited by floods. When the
floods are not very violent or very frequent, it is common for other plants from
surrounding plant communities (forest species, mainly) to appear in these
streambeds.
The
thickness of the alluvial deposits was highly variable, as was the size of the
rock fragments of which they were composed, both characteristics being related
to the slope and, in general, the orographic and lithological configuration of
the site. In some cases, the “alluvial MSS” was no more than 50 cm deep before
hitting the bedrock, whilst in other cases its depth exceeded one or one and a
half metres (Figure
4).
As
already mentioned, the loss of many of the data loggers makes it impossible to
generalise the results of the temperature and humidity parameters. Furthermore,
the measurements taken corresponded to a depth of between approximately 20 and
30 cm, and it is well known that the physical conditions of temperature and
humidity in the substratum change rapidly the further down one goes [77].
However, we can conclude that the “alluvial MSS” is an extremely damp
environment, with a RH which is almost constantly in the vicinity of saturation
levels. It should be mentioned, however, that the plastic hood in which the
data loggers were placed may have produced an overestimation of the RH due to
the condensation of water inside. Compared with the external environment, the
“alluvial MSS” is very stable as regards temperature [59].
Although the temperature in the MSS showed a pattern (cycle) similar to that of
the external environment [59,],
values for the mean daily variations in temperature above ground ranged between
10.79-14.78°C, whereas at 20-30 cm deep in the MSS, they ranged between 1.12
and 4.96°C, representing an average of 10°C lower (Figure
3 and Table
2). This is not surprising if we consider that below 50 cm in the
substratum, daily temperature variations are usually practically negligible [77],
although it may be assumed that in MSS without soil cover, such as the
“alluvial MSS”, this depth must be greater [78].
The difference in values between the external environment and the MSS for Tmax
and Tmin was around 15°C and 5°C on average, respectively. These data show that
although there was an appreciable temperature cycle, the MSS had a more modest
variation range than the external environment, and suggest that the MSS was a
more effective buffer against maximum temperatures than against minimum
temperatures [79,].
Therefore,
the “alluvial MSS”, is not only similar in structure to other MSSs (a network
of subterranean spaces among rocks and stones), but also in its degree of
isolation from surface. However, it has some particularities that differentiate
it from other MSSs. The origin of the deposit of stones is alluvial, and the
isolation from the surface is not due to a soil layer, but to a layer of silt,
sand and gravel (Figure
4b, c, d), and it may eventually have disturbances due to floods of
variable intensity.
Ecological importance of the “alluvial MSS”
It is
well known that plant and animal species associated with temporary watercourses
in arid and semi-arid regions have developed adaptive strategies that allow
them to survive water stress [81,].
Watercourses subject to natural spatial and temporal environmental disturbances
have been described by Margalef [82]
as dynamic evolutionary elements that favour the dynamic processes of species
colonisation and expansion [,]. In this paper, we show that this happens not
only on the surface and banks of the fluvial network (which includes the
edaphic environment), but also in the hypogean environment closely associated
with alluvial deposits. In line with the above, the alluvial plains subject to
periodic flooding are increasingly being studied as a whole, rather than from
an exclusively terrestrial or aquatic perspective [87]
As with the “alluvial MSS”, this periodic flooding constitutes a “pulse” that
disrupts the dynamic equilibrium these systems had reached before the flood. It
is precisely because of these disturbances that it seems difficult to conceive
of the existence of terrestrial fauna living in the “alluvial MSS” when,
periodically, these channels are completely flooded (Figure
5c-f). However, these apparently catastrophic episodes do not appear to
represent an obstacle to the survival of subterranean communities, whose
members show very different degrees of adaptation to hypogean life. The proof
of this lies in the terrestrial life forms collected in the MSS only days after
heavy rainfall. The high speed at which the water flows in the upper and middle
sections of these channels may facilitate the retention of large pockets of air
within the spaces inside the “alluvial MSS”, and thus, the survival of the
terrestrial fauna. The high carrying capacity of these watercourses suggests
the possibility that hydrochory events could occur, “seeding specimens” in
other sections of the channel suitable for hypogean life [88].
The
fauna results obtained show that despite the structural instability of this
MSS, the underground spaces in the area studied constituted a habitat that was
sufficiently suitable for colonisation both by exclusively hypogean fauna
(species only known caves in the area) and frequently hypogean fauna
(“troglophile” species sensu Sket [89]).
This habitat also acts as a “refuge” for epiedaphic, hygrophilous fauna that
are capable of occupying these spaces when necessary to shelter from the
seasonal fluctuations in temperature and humidity. Therefore, this subterranean
environment must be considered an important habitat due to the role it plays in
the conservation of fauna. Furthermore, it represents a new source of
information for determining the real biodiversity of sites which, until now,
had only been explored above ground and provides data which contribute to
improving knowledge of the real distribution of hypogean and epigean species.
Lastly, the possibility must also be raised that this new type of MSS may act
as an ecological corridor for both hypogean and epiedaphic, hygrophilous fauna.
Synthesis of characteristics of the “alluvial
MSS”
The
“alluvial MSS” is a type of terrestrial shallow subterranean habitat that
consists of a network of spaces which forms among the alluvial deposits of
temporary watercourses, and which may be covered or not by a more or less
evolved soil on which some herbaceous or woody plants may manage to grow.
Structure.
the
“alluvial MSS” forms within alluvial deposits of any type of lithology, and has
a thickness which ranges from a few decimetres to several metres deep,
overlying the bedrock. Among the gravel (a few millimetres in diameter) and
rocks (of up to several decimetres in diameter) that comprise this habitat,
empty micro and mesovoids can form which facilitate colonisation by, or provide
refuge for, terrestrial fauna presenting very diverse ecological preferences.
Origin.
this
habitat is formed as the result of the accumulation of eroded rock fragments
(principally pebbles) in dry watercourses, which have been carried there by
periodic flooding of the channels.
Abiotic factors.
it
moderates the temperature with respect to the external environment, presents
high humidity and, below a certain depth, constitutes an aphotic environment.
This habitat is usually affected by sporadic, short-lived and temporary
flooding.
Energy Sources.
nutrients
(organic detritus of animal and vegetable origin) transported by floodwater or
from the banks by surface runoff, waste products resulting from the metabolism
of the animals inhabiting the MSS and plant remains from vegetation growing on
the banks and plants growing in the channel itself (grasses and shrubs).
Fauna.
animals,
generally invertebrates and principally arthropods, of different taxa and
diverse ecological roles, including epiedaphic, sub-lapidicole, hygrophilous,
subterranean (endogean and even hypogean) and ripicolous species. During
flooding, it may also contain species which are typically aquatic.
Evolution.
this
subterranean environment appears to be more dynamic and unstable than other
types of MSS. Floods encourage the periodic creation and destruction of this
habitat’s structure, and it is the uppermost layers of the alluvial deposit
which are the most unstable. In addition, the alluvial deposit may also be affected
by two different processes that are closely related to the transportation of
sediment by water: a periodical washing away of sediments or a process whereby the
interstices of the alluvial deposit become filled with sediment.
El hábitat del que habla la noticia no es nuevo, ya que había sido previamente descubierto, descrito y comentado por Zaballos (1997): puede descargar el artículo original en:
ResponEliminahttps://www.researchgate.net/publication/258151353_Un_nuevo_Geocharis_de_Almera_(Coleoptera_Caraboidea_Trechidae_Anillini)?ev=prf_pub
Los autores del nuevo artículo han vulnerado los principios de autoridad y prioridad en los que se basan los descubrimientos científicos al ignorar el trabajo publicado 16 años antes.
En el artículo de 1997 no se habla de fauna hipogea, ni se mencionan las diversas tipologías de MSS, ni se adscribe este medio a un MSS. Se menciona la captura casual de fauna ENDOGEA en un lecho de un barranco. No obstante, Ortuño et al citan en antecedentes más trabajos en los que ha habido capturas puntuales de fauna HIPOGEA (como es la propia del MSS) en otros lechos de cauces temporales, (como Trechini adaptados, en Bucciarelli 1960, tal y como describen Giachino y Vailati 2010) y es en este trabajo donde se ADSCRIBE este medio a la tipología MSS. Es decir, no se trata de la primera vez que se citan estas capturas, ya que en el trabajo SE RECONOCEN LOS ANTECEDENTES DE FAUNA HIPOGEA CAPTURADA EN LECHOS. En este trabajo se describe una tipología de MSS, al adscribir los lechos de barrancos (y NO TODOS LOS LECHOS, SOLO LOS QUE NO SON "TERROSOS" O COLMATADOS) a los Medios Subterráneos Superficiales.
ResponEliminaEn el trabajo de Zaballos de 1997 se captura fauna con aparatos de Berlese, que se utilizan para medio ENDOGEO o EDÁFICO, y no para HIPOGEO como es el MSS. El hábitat aquí descrito (en Ortuño et al) son los espacios en un conjunto de piedras, que no se pueden poner en un aparato de Berlese.
En resumen, hablar de "fauna de lecho" dista mucho de hablar de fauna hipogea adaptada a MSS aluviales, sobre todo cuando la fauna encontrada en estos MSS aluviales es compartida con otros MSS (de tipo coluvial).
De hecho, pese a la calidad de las fotografías del trabajo de Zaballos de 1997, se puede ver que los lechos son terrosos, y estarán bastante colmatados como para albergar los espacios que necesita la fauna hipogea.
En resumen, la crítica del anterior comentario está infundada, ya que la fauna es diferente, el tipo de ambiente es diferente, y en ningún momento se habla de otros MSS.
José D. Gilgado
La publicación a la que hace referencia Zaballos, es simplemente un artículo taxonómico en el que describe una nueva especie de Geocharis capturada a partir de una muestra terrosa en el cauce de un barranco. La evidencia de fauna sobre los cauces o debajo de piedras en estos medios tiene una larga historia, otra cosa bien distinta es que se hayan estudiado adecuadamente estos medios. Es pertinente decir que los Geocharis (género de vida endogea) viven en los microespacios de medios terrosos o arcillosos, y en el caso de la muestra que se colectó en Almería (en 1997) en un lecho colmatado. Precisamente ese tipo de cauces fueron rechazados para nuestro estudio porque no alberga medio hipogeo. Puede que sirva para la vida de la fauna endogea (Geocharis y otros géneros) pero desde luego no tiene nada que ver con lo publicado por nosotros. Por ello, resulta de todo punto inaceptable que Zaballos ahora se pretenda atribuirse el mérito del descubrimiento de este hábitat de fauna hipogea, siendo aún mucho más grave que se nos acuse a los autores de vulnerar los principios de autoridad y prioridad.
ResponEliminaLa ciencia se construye sumando y no con guerras intestinas sin fundamento alguno.
Vicente M. Ortuño
Tengo poco que añadir a las aclaraciones sabiamente efectuadas por mis colegas.
ResponEliminaEs lamentable tener que soportar este tipo de acusaciones cuando,
explícitamente en el texto, se reconoce que fueron los antecedentes de capturas
de hipogeos (que no endogeos) en los lechos - con sus correspondientes referencias - los que
dieron la pista para efectuar el estudio. Estas referencias son, por cierto, anteriores
a 1997. Para más inri, el trabajo de Zaballos de 1997 NO DESCRIBE NINGÚN MEDIO SUBTERRÁNEO SUPERFICIAL (MSS).
En cualquier caso, la lectura de los dos trabajos (que cualquiera puede consultar de manera
gratuita) deja claras las diferencias y la falta de sustento del argumento del Dr. Zaballos.
Acusaciones gratuitas pero extremadamente serias que manejaremos con la contundencia que requieren.
Alberto Jiménez Valverde
Seré muy breve para dejar las cosas claras. Somos cientos los investigadores que nos dedicamos, en todo el mundo, al estudio del medio hipogeo. Esta comunidad científica ha dispuesto de 16 años para valorar la contribución a la que alude Zaballos, y no se conoce referencia alguna que reconozca la caracterización de un nuevo hábitat hipogeo, ni tampoco la referencia a fauna hipogea, en el comentario que él vierte en su artículo. A las pruebas me remito. Ahí están los hechos.
ResponEliminaLas citas que recoge Google de su trabajo de 1997, son 6 (dos de ellas autocitas) y pueden consultarse en… http://scholar.google.es/scholar?oi=bibs&hl=es&cites=11713138448860439368
… todas ellas hacen referencia a aspectos taxonómicos y sistemáticos de Anillini.
Por otra parte, la circunstancia de que pueda haber un continuo entre medio endogeo e hipogeo no significa nada, ya que es bien sabido que entre medios vecinos existe lo que conocemos por ecotono.
Un índice de la calidad de un trabajo es su cita por otros colegas, pero lo que no sucede en el mundo científico es que un autor proteste y acuse a otro investigador, por haber empleado un criterio razonado que deja fuera de posibilidad de cita su trabajo. Ese tipo de protestas sólo se producen en ambientes de autoritarismo científico. La autoridad científica se consigue por la calidad, y claridad, de los artículos no por la protesta.
Ahora, que la comunidad científica juzgue ambos trabajos y cree su propia opinión. En todo caso, la discrepancia sobre si su artículo debía haber sido citado o no, en la reciente publicación, dista mucho de justificar la acusación que se nos hace sobre haber vulnerado los principios de autoridad y prioridad.
Vicente M. Ortuño
Estimado Sr. Ortuño, aquí no estamos hablando de la calidad de un artículo ni de las veces que ha sido citado, estamos hablando de que el artículo en cuestión está en la literatura científica desde 1997. Que es el único antecedente conocido sobre la fauna del lecho de los barrancos, que en él se especifica que ese tipo hábitat debe ser considerado como nuevo, y que ustedes no lo citan en su artículo sencillamente porque esa cita dejaría, cuanto menos en entredicho, lo del “nuevo hábitat subterráneo”. No admite discusión la originalidad del artículo de Zaballos (1997), si no la originalidad del “nuevo hábitat subterráneo” (Ortuño et. al., 2013).
EliminaParece que lo de colmatado y arenoso ha quedado claro ¿no?. Según los comentarios 2 (Gilgado) y 3 (Ortuño) “En este trabajo se describe una tipología de MSS, al adscribir los lechos de barrancos (y NO TODOS LOS LECHOS, SOLO LOS QUE NO SON ''TERROSOS'' O COLMATADOS) a los Medios Subterráneos Superficiales'. Vistos los comentarios de Zaballos (1997) en el apartado “HÁBITAT” sobre el tamaño de los guijarros (dice expresamente 'gruesos guijarros' y los 'grandes espacios' que quedan entre sí), QUEDA CLARO que NO se trata de lechos 'terrosos o colmatados' y, por lo tanto, en los comentarios 2 y 3 de Gilgado y Ortuño están asumiendo/reconociendo implícitamente que el lecho descrito por Zaballos con estas características quedaría adscrito a los medios subterráneos superficiales.
Resumiendo, si no citan el antecedente por desconocimiento, MAL, y si no lo citan conociéndolo PEOR. ¿Calidad científica? ¿Honradez profesional?.
No quiero apropiarme de méritos ajenos, pero tampoco quiero que nadie se apropie de los míos.
Sin renunciar a mi punto de vista, insisto en que estamos hablando del mismo tipo de hábitat. Y el lector de ambos artículos podrá comprobar que, pese a la defensa que hacen Ortuño et al en los comentarios precedentes, son lo mismo.
ResponElimina1.- En ambos artículos se refieren a estos lugares como “Barranco” y “Rambla”, y se trabaja en el lecho de los mismos, Zaballos en Almería y Ortuño et. al. en Alicante y Valencia.
2.- En los comentarios de los autores se trata de separar los medios hipógeo y el endógeo, pero en la introducción de su artículo (pág. 1) ellos mismos reconocen que hay “ una continuidad de vida desde la superficie del suelo a través de los diferentes horizontes edáficos y el MSS....” dejando claro que existe una transición entre ambos medios y la fauna que alberga. Igualmente, los autores, en el apartado “Descripción de un Nuevo Hábitat: The “alluvial MSS” (pág. 6) dicen literalmente sobre su fauna que “Estas especies exhiben una naturaleza geófila, habitando medios estrechamente asociados con el suelo (horizontes edáficos o diferentes hábitats epiedáficos y biotopos)”. No parece lógico que los autores citen precedentes de capturas de hipógeos en lechos de ríos o arroyos, pero ignoren capturas de otra fauna edáfica en lechos de barrancos, que es el entorno sobre el que han trabajado.
PERO LO QUE SE DISCUTE AQUÍ, NO ES EL TIPO NI LA DENOMINACIÓN DE LA FAUNA QUE HABITA EN ESTE HÁBITAT, SINO EL HÁBITAT EN SÍ.
3.- En los comentarios de los autores, se hace mucho hincapié en resaltar que la especie de Geocharis procede de un lecho colmatado o terroso, cuando en el artículo de Zaballos (1997) se especifica todo lo contrario, Pág. 177: “Aunque el aspecto del lecho era de sequía total, bajo los GRUESOS guijarros aparecen otros tapizados PARCIALMENTE por tierra y lodo, que aumentan en humedad con la profundidad, y dejan entre sí GRANDES ESPACIOS por los que pueden desplazarse sin dificultad la fauna edafobia”. Y, a pesar de la escasa calidad de la fotografía 7 de Zaballos, 1997 (en blanco y negro, pág. 177) se observan sin dificultad los gruesos guijarros (se acompaña de un tubo Eppendorf como escala) y que, sorprendentemente, es prácticamente igual a lo que se aprecia en la fig. 4B, pg. 9 de Ortuño et. al., 2013; que los autores consideran como “Alluvial MSS”.
4.- No cuestiono la calidad del artículo de Ortuño et. al. ni la denominación de MSS Aluvial a este tipo de hábitat. Merecen mi reconocimiento; pero la ciencia también pasa por reconocer de forma adecuada el trabajo realizado por otros. No es lícito que el trabajo se publicite como el descubrimiento de un NUEVO medio, cuando lo que se hace es definir y acuñar un NUEVO NOMBRE para un hábitat del que ya se tenía conocimiento previo.
5.- Para terminar, considero que los comentarios precedentes de los autores Ortuño et. al., tratan de camuflar con subterfugios el tema principal. La cuestión incontestable es que existe una referencia bibliográfica que resulta relevante para el trabajo y que ha sido omitida del mismo, y es muy significativo que una cita de estas características no figure entre los antecedentes.
La referencia de Zaballos no es la primera en citar fauna en los cauces de aguas temporales. Ortuño et al citan otros en antecedentes. Hay que abundar en que en el trabajo de Ortuño et al se citan trabajos sobre fauna HIPOGEA en cauces de aguas temporales. La aparición de fauna endogea en ambientes similares no se menciona porque no tiene nada que ver. E incluso, si se refiriera a fauna hipogea (típica de cuevas), tampoco sería la primera referencia, como ya he dicho.
ResponEliminaZaballos no 'describe' un nuevo hábitat hipogeo, sino endogeo, con fauna endogea, y muestreada con berleses, como fauna endogea que es. Este tipo de medios es diferentes a los MSS, aunque haya un ecotono entre ellos.
Grandes espacios es bastante relativo. Grandes espacios para fauna como los Geocharis no lo son en absoluto para un Platyderus, por poner un ejemplo. He ahí la diferencia que hace Howarth (1983) entre micro, meso y macrocavernas, como tres tipos de ambientes diferentes, con tipos de fauna diferentes. La primera de ellas aludiendo al medio edáfico, la segunda a red de fisuras y MSS; y la tercera a grandes cuevas.
Por todo esto lo que es 'colmatado' para un MSS no tiene por qué serlo para un medio edáfico.
Si no se ha citado el trabajo de Zaballos ha sido porque no era relevante para lo aquí expuesto, ya que se ha considerado que no trataba el mismo asunto. Somos un equipo de varios investigadores que han trabajado conjuntamente en todos los aspectos del estudio y del artículo, y cualquiera de nosotros hubiera considerado relevante citar ese trabajo, lo habríamos hecho. No obstante, al considerar, insisto, en que hablaba del tipo de medio que nosotros desechábamos para nuestro estudio, y de otra fauna diferente, no lo consideramos relevante.
José D. Gilgado
Edáfico hace referencia a suelos (del griego edafos). El suelo abarca los horizontes A, B y C.
EliminaEl MSS (Medio Subterráneo Superficial) fue descubierto y descrito por Juberthie, Delay & Bouillon (1980) del horizonte C1 del suelo (pág. 25 en Juberthie et al); luego pertenece al medio edáfico aunque Gilgado diga lo contrario. Zaballos (1997) en el resumen de la pág. 1 dice textualmente “… se detalla el HÁBITAT donde fue encontrada como un NUEVO MEDIO EDÁFICO”; es decir, perteneciente al suelo sin excluir ninguno de sus horizontes.
En esa descripción del MSS Juberthie et. al. (1980, pág. 25) dicen textualmente “ Estos elementos rocosos crean entre ellos un laberinto de fisuras o pequeños espacios de oscilan de mm a cm, a veces más, que constituyen no un medio poroso….. etc” “Estas plaquetas o guijarros no calcáreos están a menudo recubiertos de una delgada capa arcillosa, y están constantemente húmedos desde 20 o 30 cm de profundidad, incluso en zonas soleadas no protegidas por cubierta vegetal”
Zaballos (1997), Pág. 177, apartado Hábitat: “Aunque el aspecto del lecho era de sequía total, bajo los GRUESOS GUIJARROS aparecen otros tapizados PARCIALMENTE por tierra y lodo, que aumentan en humedad con la profundidad, y dejan entre sí GRANDES ESPACIOS por los que pueden desplazarse sin dificultad la fauna edafobia”. Es decir, la que vive en el suelo.
Juberthie et al (1980, Págs. 34y 35) dedican un apartado a “Ejemplos de mezcla de fauna endogea y subterránea” donde dicen que los medios subterráneos y endógeos “se diferencian esencialmente por su tipo de porosidad y por consecuencia por la talla de los huecos. El medio endogeo tiene una porosidad de intersticios y sus huecos MUY PEQUEÑAS dimensiones. El medio subterráneo tiene una porosidad de fisuras, constituyendo una red de espacios más anchos oscilando del MILÍMETRO a 10 CM de ancho”
Si tomamos estas medidas como referencia de tamaño de espacios en MSS, caben tanto Geocharis como Platyderus. Que la mayoría de las especies de Geocharis puedan moverse en espacios reducidos (microespacios) no quita para que no lo puedan hacer también en espacios más amplios tipo MSS. Tanto en el resumen, como en la descripción del hábitat se dice que Geocharis amicorum fue encontrado en él, no que necesariamente viva ni forme parte del mismo. Asumir que la captura se produjo en un medio terroso y colmatado porque el género es mayoritariamente endógeo no tiene ningún tipo de fundamento.
Juberthie et al (1980, Pág. 47), en las conclusiones refuerzan las ideas ya comentadas.
Ortuño et. al. (2013, Pág. 8): “Proponemos llamar a estos espacios subterráneos localizados en el sustrato aluvial “Aluvial MSS”, un hábitat hipogeo compuesto de grietas y fisuras en el sustrato que se forma entre la grava y GUIJARROS DE TAMAÑO VARIADO que constituyen los depósitos aluviales de CAUCES SECOS”
Zaballos (1997), Pág. 177, apartado Hábitat: “Aunque el aspecto del lecho era de SEQUÍA TOTAL, bajo los GRUESOS GUIJARROS aparecen otros tapizados parcialmente por tierra y lodo, que aumentan en humedad con la profundidad, y dejan entre sí GRANDES ESPACIOS por los que pueden desplazarse sin dificultad la fauna edafobia”.
El hábitat que se describe junto a la descripción de Geocharis amicorum es un MSS aluvial y, aunque el nombre que dan Ortuño et. al. (2013) es muy acertado, NO SUPONE UNA NOVEDAD. La cita de Zaballos (1997) ignorada voluntariamente (comentario nº 12 de Gilgado “Si no se ha citado el trabajo de Zaballos ha sido porque no era relevante para lo aquí expuesto, ya que se ha considerado que no trataba el mismo asunto.”) debería haber figurado INEXCUSABLEMENTE en este trabajo.